






苹果果肉硬度很大程度上决定了其采后储存、运输的便利性,也是消费者选择的一个重要因素,同时也影响苹果自身种子繁殖,因此阐明苹果果肉硬度的遗传调控机制,对苹果育种具有重要的实际意义。
近日,西北农林科技大学赵政阳研究团队在Plant Physiology上发表了题为“Upregulation of PECTATE LYASE5 by a NAC transcription factor promotes fruit softening in apple”的研究论文,该研究发现果肉硬度受多基因调控,通过BSA、重测序等技术筛选到了影响果肉硬度的关键基因MdPL5,并发现MdPL5基因上游的转座子插入可以抑制其表达,影响苹果硬度;并利用酵母单杂交、RNA-seq等筛选到了MdPL5基因的上游调控因子MdNAC1,并通过遗传转化实验进行了基因功能表征,确定了其在果肉软化中的作用。
本项目中所用的苹果酵母cDNA文库由欧易生物构建。

文库与诱饵
酵母文库:苹果酵母cDNA文库
诱饵基因启动子:MdPL5-promoter(542 bp)
筛选方法:酵母单杂交(Y1HGold-pAbAi系统)
验证方法:Y1H assay/EMSA/dual-LUC/ChIP-qPCR
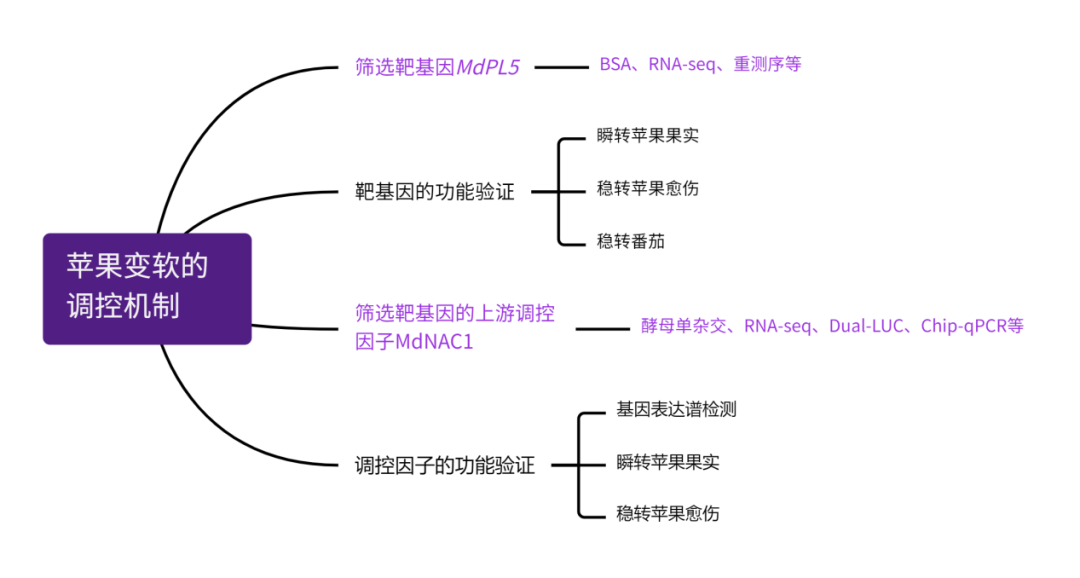
1、F1代群体中苹果果肉硬度的QTL鉴定
前期有研究将MdACS1和MdACO1作为苹果果肉硬度的分子筛选标记。然而本研究在构建的439个“粉红”和“富士”杂交的F1代杂交苹果中发现,MdACO1基因型在极端杂种中没有出现分离现象;在苹果收获时,MdACS1标记的基因型也与果肉硬度无明显相关性。因此,研究者推测可能存在其他的分子标记和调控机制影响成熟期果肉硬度。
研究者在F1代中选取了果肉超硬和超软2组苹果,每组各20个,进行BSA测序,结果分析获得了10个QTL区域,共包含1109个基因(图1E)。结合超硬和超软苹果果肉的比较转录组测序数据(图2A),发现存在84个DEGs,其中7个与细胞壁降解发育有关。进一步结合基因组重测序结果(样本同BSA-seq)分析基因组上的遗传变异位点,发现了一个2432-bp的转座子SV,位于果胶酸裂解酶基因MdPL5(MD16G1106400)转录起始位点上游4.7kb,命名为TE-2432。
研究结果显示,与超硬果肉组相比,该SV片段的缺失频率在超软组中更普遍(图2B,C),纯和缺失基因型的果肉硬度显著低于del-2432基因型的苹果果肉(图2D),在超软果实组中明显的纯合子缺失基因型(Del-2432,aa)缺失率为66.7%(图2E)。这些结果表明,TE-2432与苹果收获时的果肉硬度有较明显的连锁关系,TE-2432可作为预测成熟时果肉硬度的分子标记,邻近的MdPL5可能是一个很有前途的果实硬度候选基因,值得通过功能验证进行进一步研究。
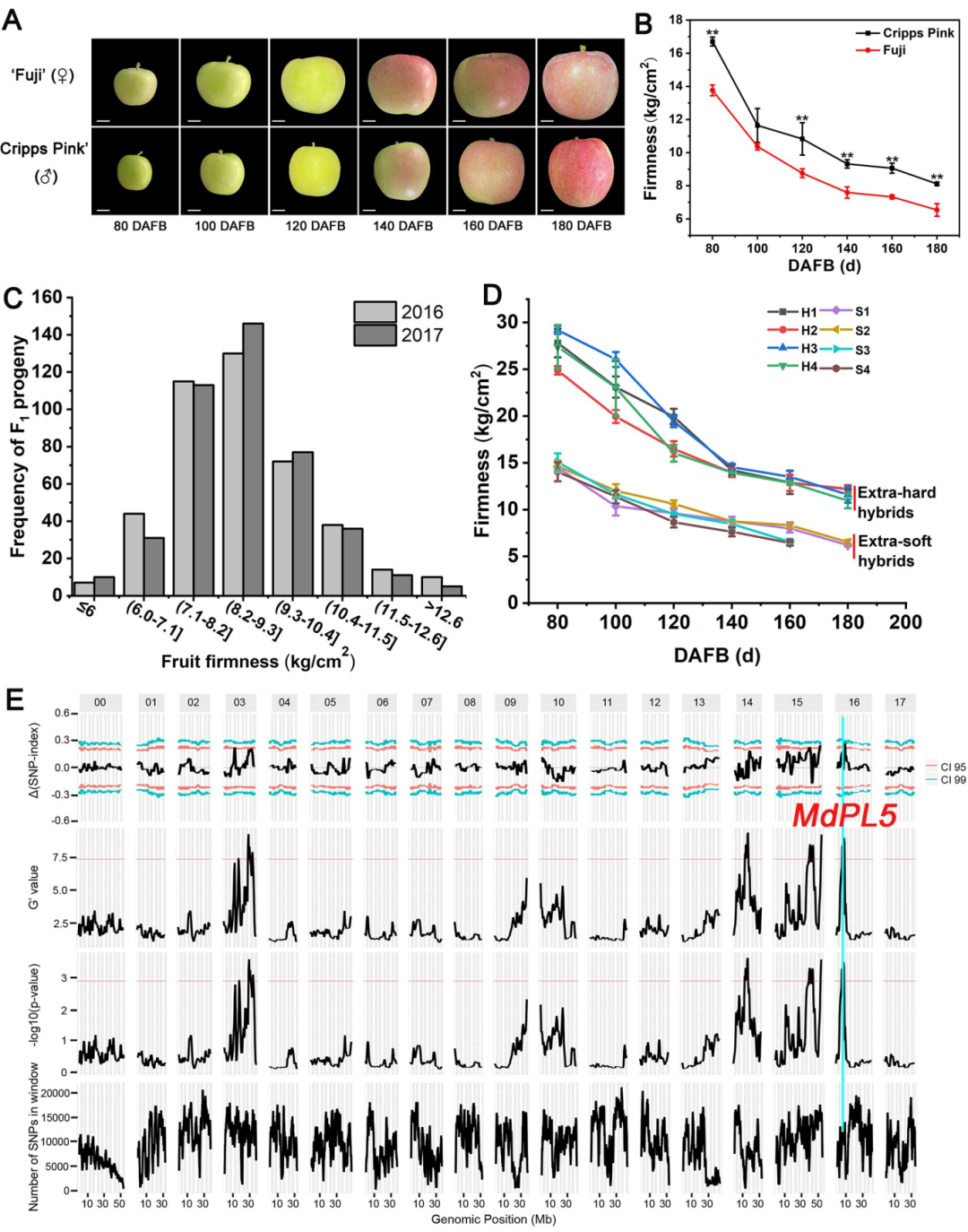
图1 |“富士”和“克里普斯粉红”F1群体果肉硬度的QTL鉴定
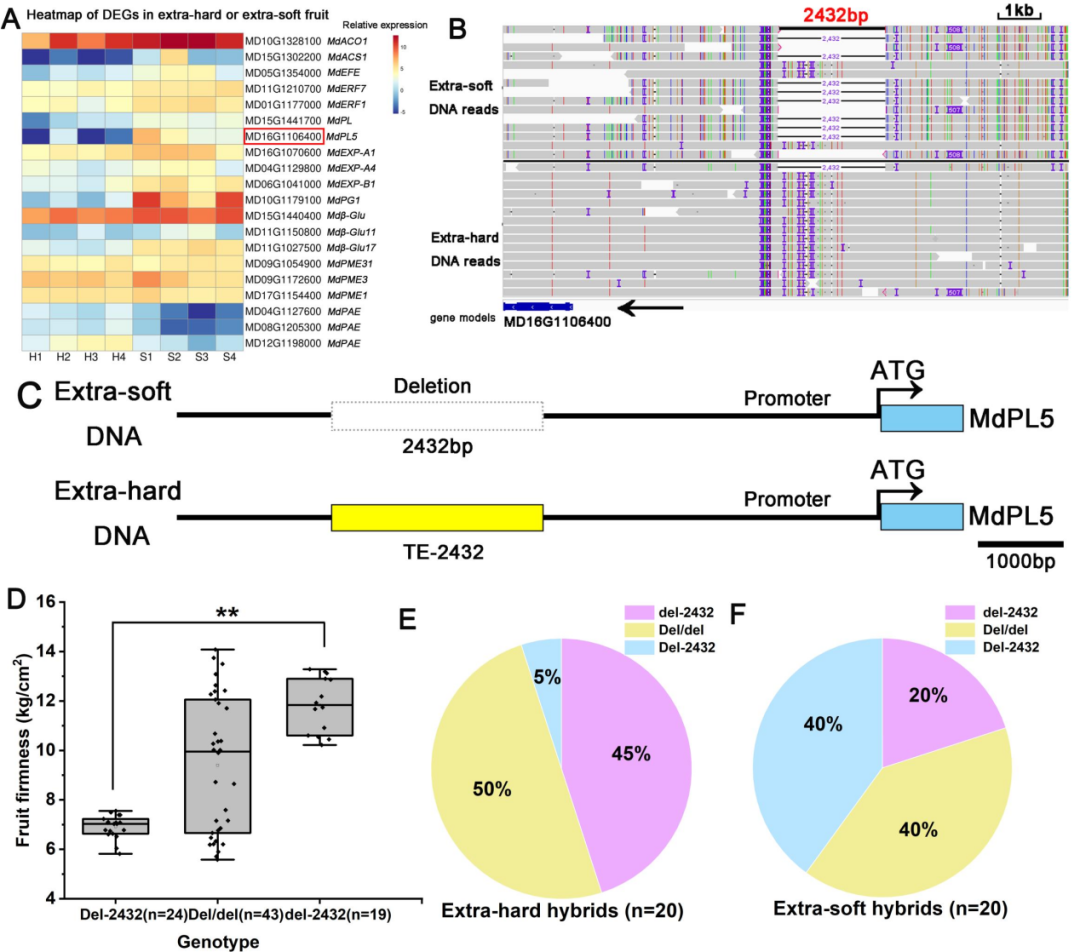
图2 | F1代果肉硬度调控的候选基因鉴定
2、MdPL5基因的表达特征及对果肉硬度的功能表征
研究者进一步对MdPL5基因的表达谱和功能表征进行了检测,结果发现其主要在花和果肉中表达,在“富士”中的表达量显著高于“粉红”(图3AB),F1代中超软果肉的表达量显著高于超硬组(图3DF),说明MdPL5对果肉软化可能起正调控作用。此外,通过LUC检测发现2432-bp转座子的插入,对MdPL5的表达起抑制作用(图3GH)。
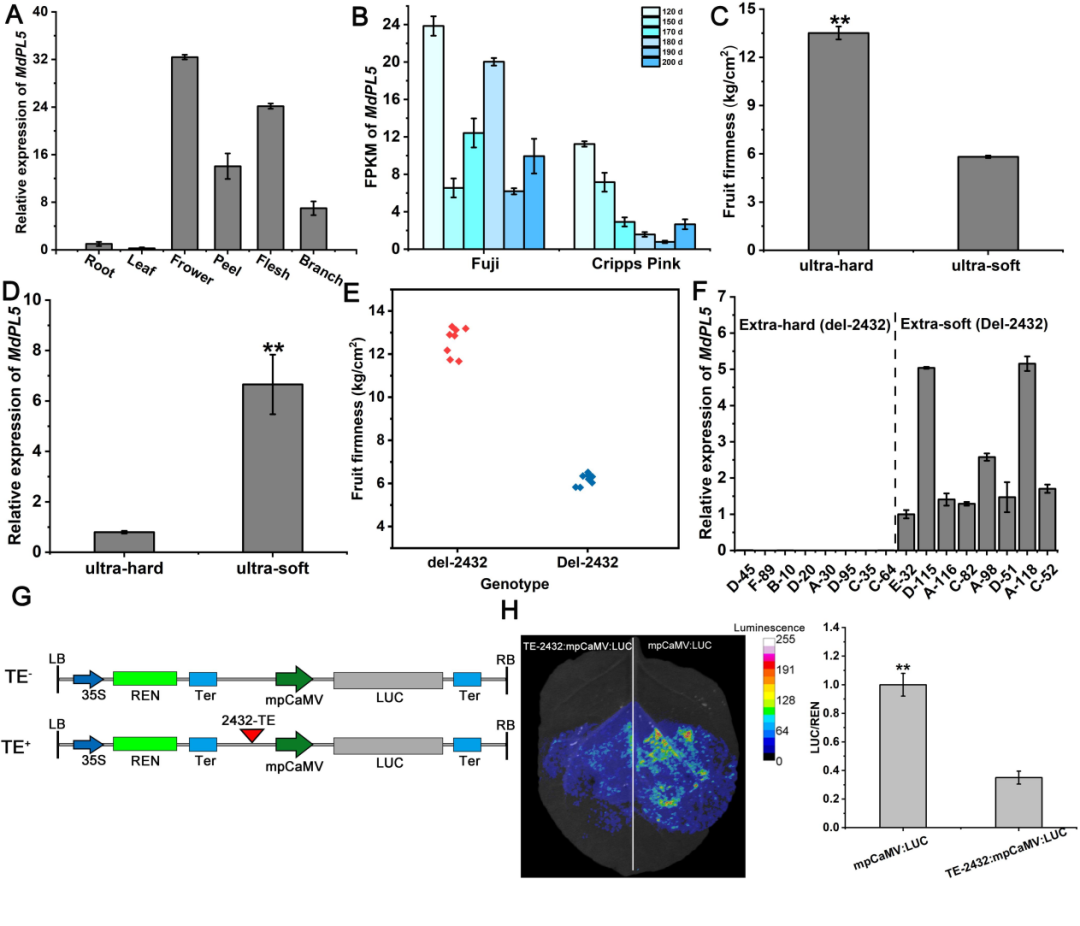
图3 | MdPL5基因的表达谱鉴定
接着,研究者从苹果中克隆了MdPL5基因,并构建了植物过表达载体MdPL5-pCAMBIA2300和基因沉默载体MdPL5-pTRV2。在苹果果实瞬转结果中发现,过表达5天的苹果MdPL5表达量显著增加,果实硬度降低;沉默表达的苹果没有明显的表型差异(图4A-C)。在生理指标上,过表达苹果PL、PG和β-gal酶的活性显著提高,而沉默MdPL5则抑制了PL和PG酶的活性(图4D-F)。在稳转苹果愈伤结果中发现,与对照组相比,MdPL5在愈伤组织中的过表达并没有导致明显的表型变化(图5A)。但水溶性果胶(WSP)、螯合剂可溶性果胶(CSP)、PL、PL、PG和β-gal酶的活性均较对照组显著升高,而碳酸钠可溶性果胶(SSP)的含量急剧下降(图5D-I)。
此外,研究者还在番茄中对MdPL5进行了功能验证,同样发现过表达MdPL5对番茄发育及收获后的软化具有促进作用,且与对照组相比,番茄成熟时间提前了,但果实大小不变(图6)。

图4 |MdPL5正向调节苹果果肉软化
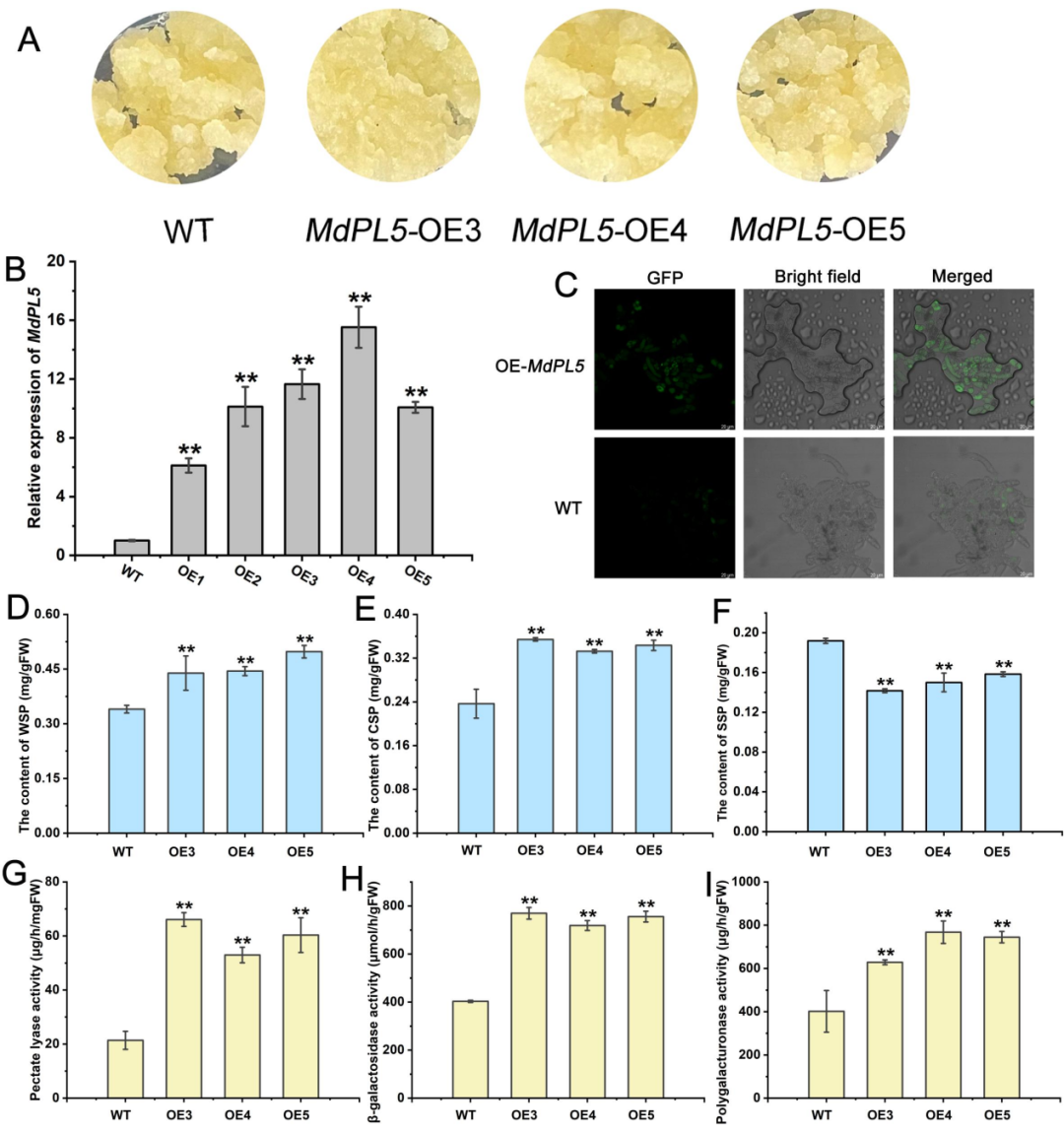
图5 | MdPL5在转基因苹果愈伤中的功能验证
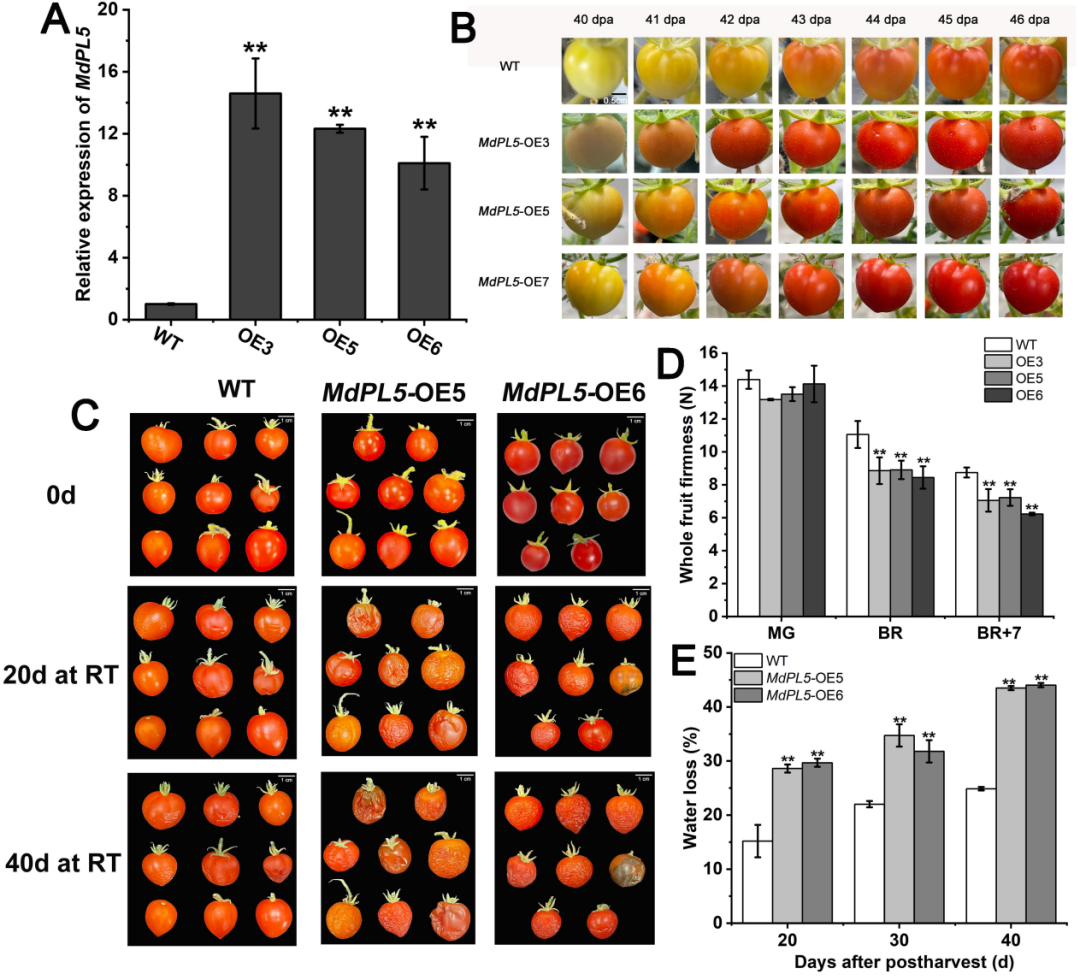
图6 | MdPL5对转基因番茄果实发育及采后软化的功能验证
3、MdNAC1-L调控MdPL5的转录表达
前面研究发现2432-TE的纯和插入会抑制MdPL5的表达,然而在同一品种苹果的不同发育时期MdPL5的表达水平也存在显著差异(图3B),因此研究者推测可能还有其他的机制调控MdPL5的转录。逐个排除了CDS可变剪切、启动子活性差异的影响因素,最终通过酵母单杂交文库筛选及RNA-seq,找到了调控MdPL5表达的上游转录因子MD03G1210100、MD16G1069500、MD16G1185900、 MD05G1361500。其中MdNAC1-L (MD16G1069500)的表达量随着苹果果实的发育时间逐步升高,认为可能与MdPL5调控果实软化有关,被选择继续进行了一对一体内外互作验证,结果证实MdNAC1-L确实与MdPL5启动子互作并调控其转录(图7)。
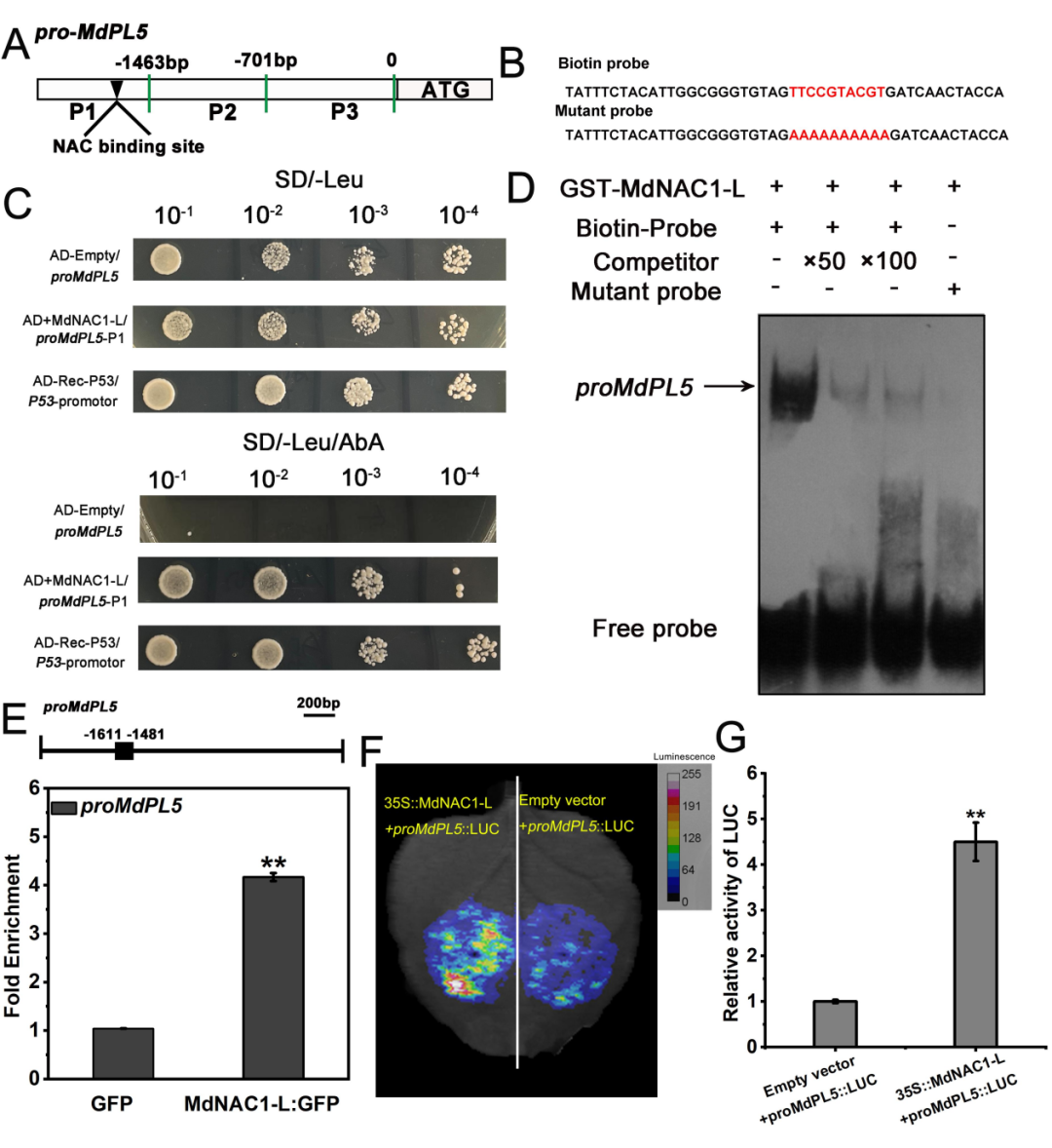
图7 | MdNAC1-L结合MdPL5启动子并促进其转录活性
4、MdNAC1-L的表达特征及加快苹果果肉软化的功能表征
研究者进一步对MdNAC1-L基因的表达特征进行了检测,其中系统进化发现在不同物种中MdNAC1-L具有较高的保守性,与梨最同源(图8A)。随着果实发育时间延长,表达量逐渐升高,且超软苹果中的表达量高于同期超硬苹果(图8D)。亚细胞定位显示定位在细胞核,酵母自激活检测发现MdNAC1-L具有转录激活活性,说明其为典型的转录因子(图8)。
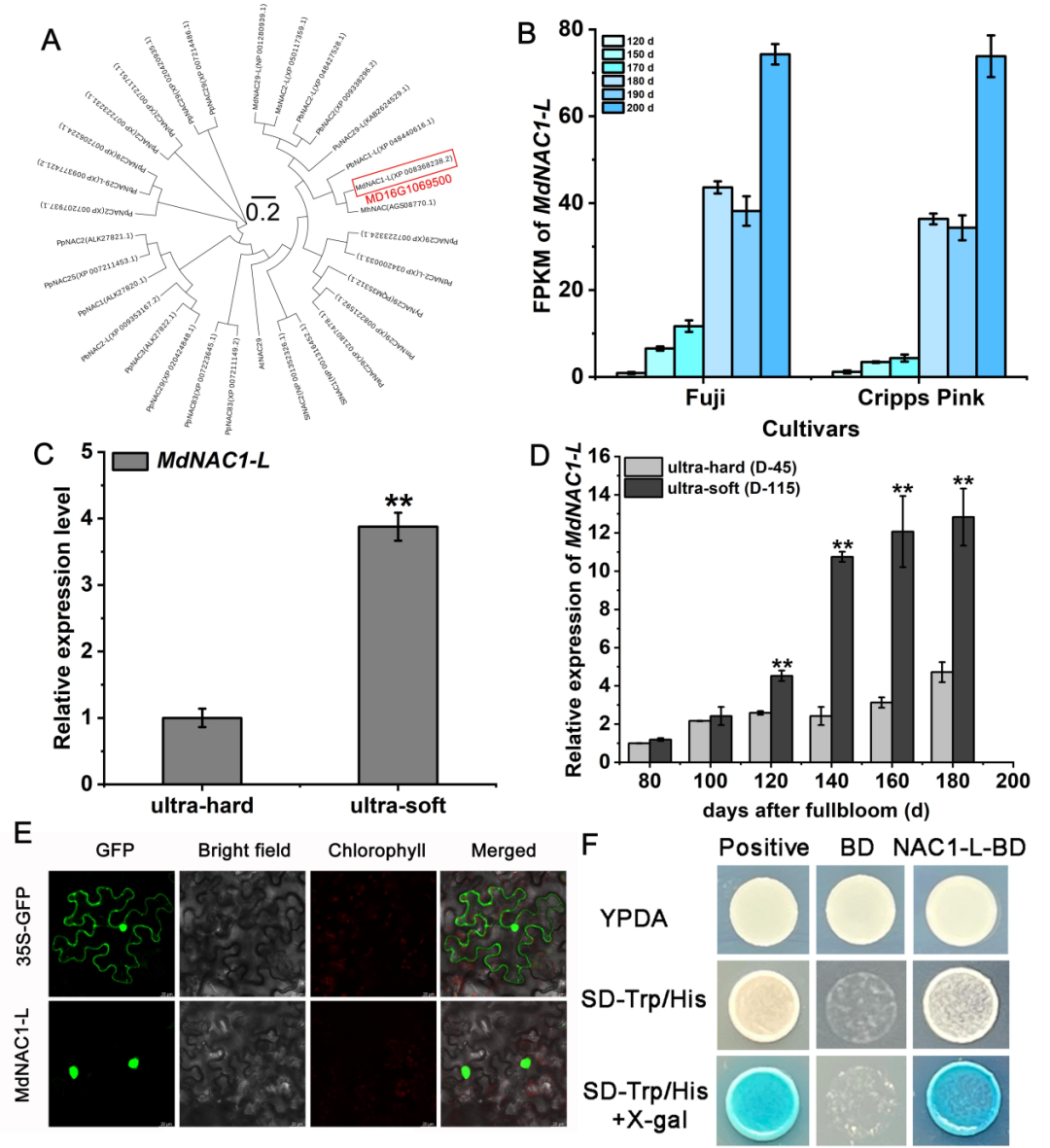
图8 | MdNAC1-L基因的表达特征及转录活性鉴定
基于以上数据,研究者也对MdNAC1-L通过瞬转苹果果实和稳转苹果愈伤,进行了果实成熟及软化方面的功能表征研究。在瞬转苹果果实中发现MdNAC1-L增强了果实软化进程,过表达组果肉细胞完整性显著低于MdNAC1-L沉默组。在稳转苹果愈伤中还发现,MdNAC1-L过表达组中果胶降解酶等显著升高,乙烯合成基因MdACS1、MdACO1和果胶相关基因MdPG1的转录量显著提高。这些数据表明,MdNAC1-L不仅正向激活乙烯合成基因的表达,促进乙烯的产生,而且还直接调节果胶降解酶,从而诱导果实软化(图9)。
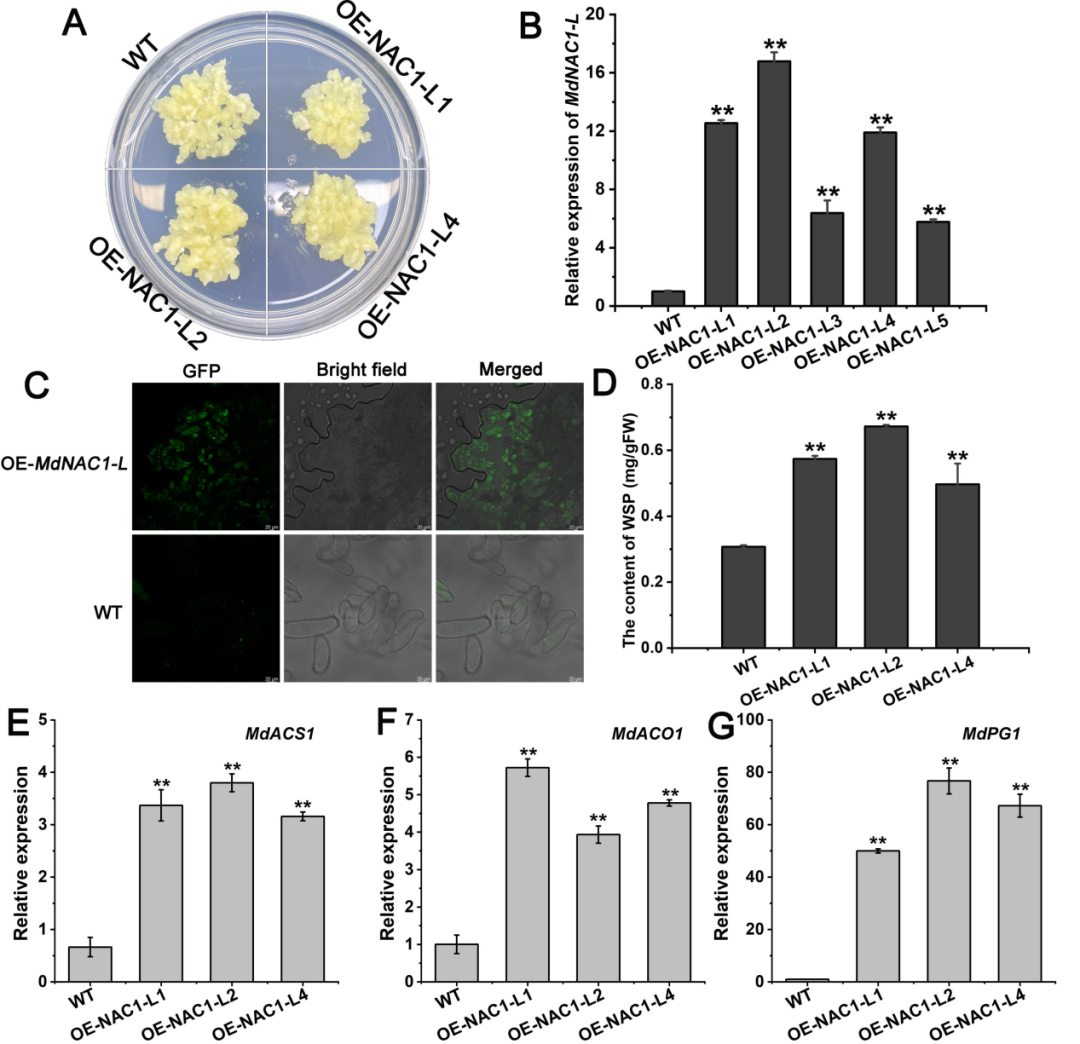
图9 | MdNAC1-L在稳定的转基因苹果愈伤组织中的功能验证
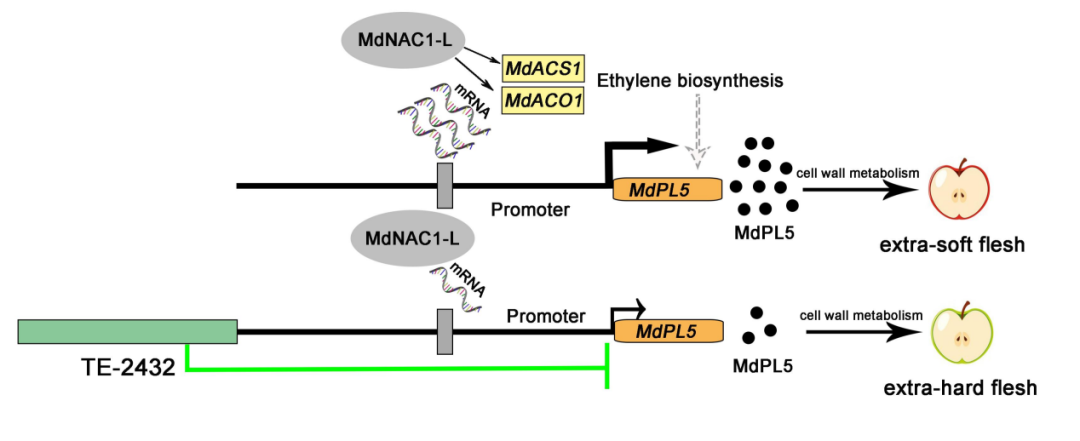
图10 | TE-2432&MdNAC1-L&MdPL5调控苹果果实硬度的工作模型
本研究通过BSA-seq等发现了新的苹果果肉硬度调控QTL,包括MdPL5基因以及能够调控其转录的上游转座子2432-TE,并通过酵母单杂交筛选、RNA-seq及一系列遗传转化等实验发现MdNAC1-L作为转录激活因子诱导MdPL5及乙烯相关基因的表达,从而调控果实成熟和软化。
西北农林科技大学园艺学院赵政阳教授为本文通讯作者,博士研究生苏秋方和青年教师杨惠娟为论文共同第一作者。该研究得到国家自然科学基金项目和国家现代农业产业技术体系建设专项资金的资助,西北农林科技大学园艺科学研究中心为本研究提供了部分技术支持。
DOI:https://doi.org/10.1093/plphys/kiae428