






2025年12月24日,徐州医科大学附属医院吕凌教授与南京医科大学第一附属医院的古鉴教授研究团队相关研究成果以“Tumor-produced ammonia is metabolized by regulatory T cells to further impede anti-tumor immunity”为题,被国际顶级期刊Cell接收发表。作者通过空间多组学系统揭示Tregs细胞的代谢特征,研究了肿瘤来源氨被Tregs细胞代谢利用,从而进一步抑制抗肿瘤免疫并促进免疫治疗耐药的关键机制。
作者使用10x Genomics Visium CytAssist平台进行空间转录组检测,对相邻切片通过AFA-DESI平台(Beijing Victor Technology Co., LTD,Beijing,China)与QE-HF Orbitrap质谱仪(Thermo Scientific,U.S.A)进行空间代谢组检测。
【欧易生物为本研究提供空间代谢组、空间转录组、转录组技术服务】
调控性T细胞(Tregs)如何适应恶劣的肿瘤代谢微环境以抑制抗肿瘤免疫的具体机制,目前尚不完全明确。本研究通过空间多组学分析发现,人类肝细胞癌组织中存在以高谷氨酰胺分解-低尿素循环为特征的代谢异质性区域,该区域内Tregs广泛分布,而CD8+及CD4+效应T细胞则出现凋亡。作者发现Tregs细胞通过上调精氨琥珀酸裂解酶(ASL)表达,利用尿素循环实现氨解毒;同时,氨也可在FOXP3转录因子调控的精胺合酶(SMS)作用下转化为精胺。X射线晶体学分析证实了精胺与PPARγ之间的直接相互作用,该作用可全面调控多种线粒体复合体蛋白的转录,从而增强Tregs细胞的氧化磷酸化能力及免疫抑制功能。临床观察显示,经抗PD-1治疗后死亡的肿瘤细胞通过转脱氨作用释放氨,进而强化Tregs细胞功能并导致免疫治疗耐药。针对氨生成过程以抑制Tregs细胞功能,可能为抗肿瘤免疫治疗提供一种潜在策略。

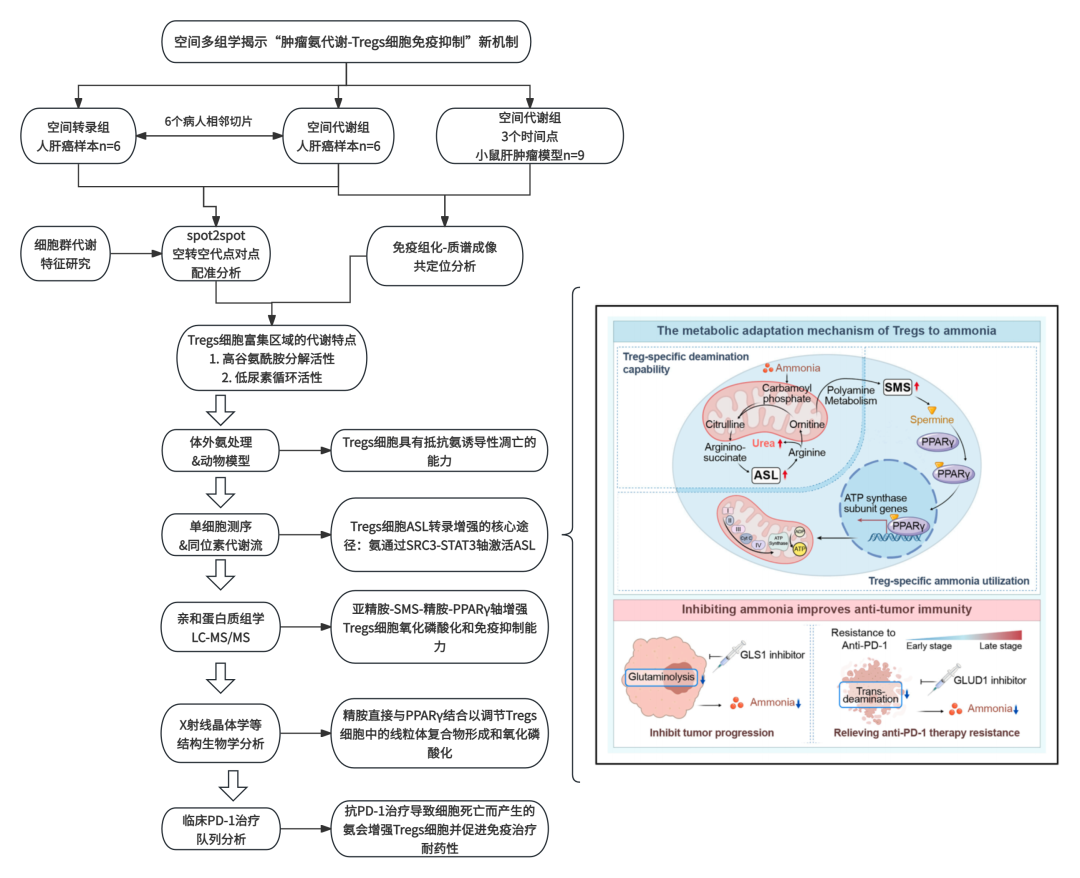
作者采用“时空多组学”新技术,揭示了Tregs细胞氨代谢“变废为宝”的概念。采用spot2spot点对点配准的创新型分析方式,整合空间转录组和空间代谢组检测数据,首次揭示氨不仅是代谢废物,更是Tregs细胞的特定“养料”和信号分子。文章具备亮眼的分子机制网络探究,发现Tregs细胞应对氨的双重机制:一方面通过ASL/尿素循环“解毒”,另一方面通过合成精胺“利用”氨来供能,促进氧化磷酸化。同时,也采用了结构生物学新方法,利用X射线晶体学等技术诠释了精胺直接结合核受体PPARγ的结构基础,这在代谢免疫领域非常深入。立足于临床耐药新机制以研究,提出了免疫治疗杀伤肿瘤后的“副作用”以及干预新靶点,具备临床转化意义。
组织样本: 人肝癌样本(6位病人,空转空代各6例,为相邻切片)、小鼠肝癌样本(3个时间点9例空代)
1. Tregs富集肿瘤亚区的关键代谢特征:高谷氨酰胺分解和低尿素循环活性
作者使用空间转录组进行免疫细胞注释,确定T细胞亚型分布微区,其中Tregs细胞在肿瘤组织内分布明显,其他亚型主要分布肿瘤周边和临近正常组织中(图 1C、B)。
作者使用上海欧易生物开发的spot2spot点对点配准的专利技术,创新性的将空间转录组条形码空间信息和空间代谢组像素点信息转换为统一的空间标识符,整合了空间转录组和空间代谢组数据,该方法旨在研究细胞群的代谢特征。
对Tregs细胞富集区域进行了代谢水平的进一步研究,作者发现:谷氨酰胺分解代谢是Tregs富集亚区的独特代谢特征(图 1D、E)。为验证这一发现,作者在9例样本,包含3个阶段的小鼠肝肿瘤模型中进行了空间代谢组检测。作者采用将荧光信号转换丰度特征结合空间代谢成像特征分析这一新颖的方法,进行了免疫组化图片和代谢物的空间共定位分析。免疫荧光结果证实,随疾病进展,Tregs细胞丰度和谷氨酸代谢活性显著增加(图 S1)。上述实验结果证明了肿瘤微环境中,谷氨酰胺分解代谢与Tregs细胞存在空间相关性。

图1. 高谷氨酰胺分解和低尿素循环活性是Tregs富集亚区的关键特征
作者进一步采用了代谢集功能变异分析的方法评估了谷氨酰胺分解代谢活跃区域中的免疫细胞组成,并定位到了Tregs细胞定位的另一要素:低尿素循环活性。高谷氨酰胺分解和低尿素循环活性的组合特征综合决定了Tregs细胞的定位(图 1F-H、图S1)。
氨是谷氨酰胺分解的主要副产物,主要通过尿素循环进行处理,作者推测肝脏相关疾病可能诱导氨蓄积,从而形成免疫抑制微环境。

图S1. 免疫组化结合小鼠模型空间代谢组验证
2. Tregs细胞的独特优势:具有抵抗氨诱导性凋亡的能力
为探究氨积累驱动Tregs细胞浸润,作者从健康人外周血分离出Tregs细胞和其他T细胞亚型,并置于不同浓度的氨环境中。流式细胞术和蛋白印迹实验结合小鼠体内实验,进一步证明,低氨条件下,Tregs维持稳定细胞内氨水平,相对其他T细胞亚型,Tregs更能抵抗细胞凋亡。氨可选择性诱导CD4+和CD8+T细胞凋亡,同时维持Tregs细胞存活(图 2)。

图2. Tregs可抵抗氨诱导的细胞凋亡;CD8+或CD4+ T细胞则不具备此抗性
3. Tregs细胞氨代谢的关键开关:精氨基琥珀酸裂解酶(ASL)
为进一步阐释Tregs细胞相对于其他T细胞亚型的氨代谢优势,作者从人类肝细胞癌(HCC)中分离出各种T细胞亚型,Tregs细胞氨水平较低并具有丰富的尿素循环中间产物(图 3A、B)。作者通过15N同位素代谢流实验结果发现,Tregs细胞尿素循环中间产物同位素丰度更高,Tregs细胞具有更强的尿素循环活性(图 3C、D)。作者使用单细胞公共数据集进一步研究发现,Tregs细胞ASL表达水平显著高于其他T细胞亚型。细胞实验和氨饮食小鼠模型结果进一步证实氨暴露通过上调ASL促进Tregs中尿素循环的激活(图 3F-H)。ASL基因敲除实验发现ASL被抑制就会导致Tregs细胞增殖能力和抗凋亡能力下降,进一步证明了精氨基琥珀酸裂解酶(ASL)是Tregs细胞氨代谢的关键开关(图 3I-M)。

图3. 氨暴露通过上调ASL表达诱导Tregs细胞中尿素循环激活
4. Tregs细胞ASL转录增强的核心途径:氨通过SRC3-STAT3轴激活ASL
通过氨处理细胞的RNA测序,ASL启动子预测分析表明,信号转导及转录激活因子3(STAT3)是Tregs细胞ASL转录的主要因素。后续STATA3免疫沉淀和质谱检测结果表明,类固醇受体共激活因子3(SRC3)是氨诱导的STAT3共激活因子中的重要因素。单细胞测序分析结果表明,SRC3的表达与Tregs细胞中ASL呈特异性正相关。后续实验表明,氨显著诱导Tregs细胞ASL转录,增强核内STAT3-SRC3复合体形成。氨促进SRC3与STAT3互作从而驱动ASL转录,增强Tregs细胞氨解毒能力。
5.氨通过FOXP3驱动SMS表达促进Tregs细胞氧化磷酸化
转录组测序显示Tregs细胞在富氨环境中氧化磷酸化(OXPHOS)显著增强(图 4A、B)。电镜结果表明,氨处理Tregs细胞线粒体保持完成,ATP生成和膜电位增加(图 4D-F)。ASL敲低降低Tregs细胞耗氧率(OCR),其过表达未恢复CD8+T细胞氧化磷酸化,表明ASL并非是氨诱导OXPHOS增加的主要因素。
肝细胞癌(HCC)和外周Tregs细胞特异性上调了与氨代谢相关的精胺合酶(SMS),液质联用(LC-MS/MS)证实肿瘤来源的Tregs细胞中精胺水平升高(图 4G -I)。SMS敲低消除了氨诱导的OCR增强和线粒体完整性,抑制细胞内精胺代谢,导致氧化磷酸化水平下降并损害了免疫抑制功能(图 4J-N)。FOXP3是Tregs细胞特异性转录因子,同时是SMS的潜在调节因子。FOXP3与SMS启动子的结合通过染色质免疫沉淀ChIP-qPCR和电泳迁移率变动分析(EMSA)得到了验证。这些结果表明FOXP3是驱动SMS表达的转录激活因子,增强Tregs细胞特异性的氨诱导的氧化磷酸化。

图4. 氨通过FOXP3驱动SMS表达促进Tregs细胞氧化磷酸化
6. 亚精胺-SMS-精胺-PPARγ轴增强Tregs细胞氧化磷酸化和免疫抑制能力
实验证明外源精胺无法进入细胞,SMS催化亚精胺转化为精胺,OCR分析表明:内源产生精胺(亚精胺转化为精胺)时,较高的细胞内亚精胺水平才会显著增强Tregs细胞的OXPHOS(图 5C)。相反,过表达精胺降解限速酶,降低亚精胺、精胺水平和OXPHOS(图 5D-F),这证实了精胺是氨存在时增强Tregs细胞OXPHOS的关键代谢物。作者使用亲和蛋白质组学,通过LC-MS/MS检测和综合分析,确定过氧化物酶体增殖物激活受体γ(PPARγ)是Tregs细胞中精胺的主要靶点(图 5H-K)。敲低PPARγ,可逆转氨诱导的OXPHOS增强(图 5P)。这一结果表明亚精胺-SMS-精胺-PPARγ轴是Tregs细胞功能增强的途径。

图5. 精胺通过PPARγ增强Tregs细胞的氧化磷酸化和免疫抑制能力
7. 精胺直接与PPARγ结合以调节Tregs细胞中的线粒体复合物形成和氧化磷酸化
为验证精胺与PPARγ结合机制,作者运用微尺度热泳(MST)、等温滴定量热法(ITC)、细胞热位移分析(CETSA)、药物亲和力响应靶标稳定性分析(DARTS)、分子对接分析、X射线晶体学等丰富的结构生物学手段,发现精胺通过静电和氢键与PPARγ的Glu319、Ser370和Glu371残基相互作用(图 6A-H)。突变这些残基为丙氨酸后,消除了精胺的结合,Tregs细胞的氧化磷酸化也无法被氨增强(图 6I-N)。

图6. 精胺直接与PPARγ结合,并调节Tregs细胞中的线粒体复合物形成和OXPHOS
8. 抗PD-1治疗导致细胞死亡而产生的氨会增强调节性T细胞并促进免疫治疗耐药性
最后,作者结合Anti-PD-1免疫治疗背景,作者发现抗PD-1治疗杀死的肿瘤细胞通过“转氨脱氨作用”(Transdeamination)释放大量氨,而非谷氨酰胺分解。这些氨进一步促进Tregs细胞增殖,导致继发性耐药。联合使用GLUD1抑制剂(R162)比GLS1抑制剂更能有效降低氨水平并克服耐药(图 7)。

图7. 抗PD-1治疗导致细胞死亡而产生的氨会增强Tregs细胞并促进免疫治疗耐药性
肿瘤浸润性Tregs细胞会适应肿瘤微环境中的代谢变化,从而形成免疫抑制性微环境,促进肿瘤进展。通过空间多组学分析,作者注释了多种T细胞亚型,并对T细胞亚型的代谢特征进行了深入研究。作者重点关注氨的生成和解毒过程,发现Tregs细胞浸润的肿瘤区域表现出增强的谷氨酰胺分解代谢和降低的尿素循环活性,这种代谢状态有利于局部氨的积累。通过一系列严谨的实验证明Tregs细胞上调ASL,以氨依赖的方式促进氨解毒。其次,多胺代谢和尿素循环中间产物,在FOXP3转录的酶SMS作用下转化为精胺,精胺与PPARγ结合,驱动氧化磷酸化上调,进一步增强Tregs细胞的功能。作者确定了Tregs细胞特异性的两种氨适应机制:SRC3/STAT3-天冬氨酸转氨酶驱动的尿素循环途径和SMS-精胺-PPARγ轴,这加深了对Tregs细胞中多胺代谢的理解,为Tregs细胞对氨或多胺的特异性反应提供了机制解释。
作者采用“时空多组学”新技术,揭示了Tregs细胞氨代谢“变废为宝”的概念。采用spot2spot点对点配准的创新型分析方式,整合空间转录组和空间代谢组检测数据,首次揭示氨不仅是代谢废物,更是Tregs细胞的特定“养料”和信号分子。文章具备亮眼的分子机制网络探究,发现Tregs细胞应对氨的双重机制:一方面通过ASL/尿素循环“解毒”,另一方面通过合成精胺“利用”氨来供能,促进氧化磷酸化。同时,也采用了结构生物学新方法, 利用X射线晶体学等技术诠释了精胺直接结合核受体PPARγ的结构基础,这在代谢免疫领域非常深入。立足于临床耐药新机制以研究,提出了免疫治疗杀伤肿瘤后的“副作用”以及干预新靶点,具备临床转化意义。
参考文献:
PMID: 41448182
DOI: 10.1016/j.cell.2025.11.034