






前言
苹果腐烂病由Cytospora mali(异名Valsa mali)真菌引起,是全球苹果产业最具毁灭性的枝干病害之一,常导致树皮溃烂、枝干枯死,严重威胁果园生产与产业可持续发展。目前高抗优质苹果种质资源稀缺,解析苹果先天免疫调控机制、挖掘关键抗病基因,成为抗病育种的核心突破口。
近日,西北农林科技大学黄丽丽、冯浩研究团队于Plant Biotechnology Journal发表的题为“TMdRLKT1–MdRAX2–MdMKS1 Module Positively Regulates Resistance to Cytospora mali in Apple”研究论文,首次阐明了受体类激酶–转录因子–下游靶基因的磷酸化级联通路,揭示苹果抵御腐烂病的全新免疫调控机制,为苹果抗病分子育种提供关键基因资源与理论支撑。本项目中受Cytospora mali侵染的苹果枝条酵母文库由欧易生物构建。

研究背景
植物先天免疫依赖细胞膜受体识别病原信号,并通过磷酸化等翻译后修饰快速传递免疫指令。前期研究发现,苹果MdRLKT1基因可被病原菌小RNA沉默以抑制宿主免疫,但其如何传递免疫信号、通过磷酸化调控下游通路尚不清楚。
本研究围绕MdRLKT1展开,系统解析其互作蛋白、磷酸化位点、下游靶基因及完整调控模块,明确其在苹果抗腐烂病中的核心功能。
文库与诱饵
酵母文库:苹果枝条(受Cytospora mali侵染)组织cDNA文库
诱饵蛋白:MdRLKT1
筛选方法:酵母双杂交(GAL4系统)
验证方法:Y2H assay/LCI/Co-IP等
技术路线
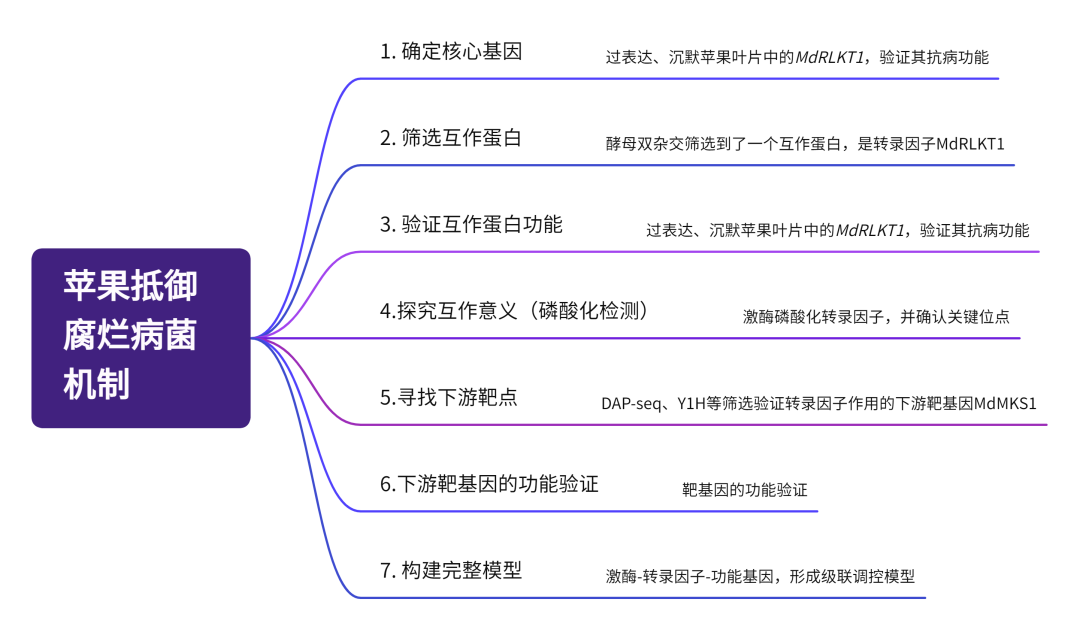
研究结果
1. MdRLKT1 是苹果抗腐烂病的正向调控因子
研究者构建了稳定抑制 MdRLKT1 表达的转基因苹果植株(Ri1 和 Ri2),发现MdRLKT1基因沉默显著降低了苹果叶片对腐烂病菌的抗性,结果显示:
·表型上,与野生型相比,MdRLKT1 表达受抑的植株表现出对腐烂病菌更高的敏感性,病斑面积明显增大(图1b);
·生理与分子响应上,MdRLKT1 表达受抑植株中的活性氧和木质素积累减少,且防御相关基因(如 MdPR1/2/4/5)及活性氧产生相关基因(MdRBOHD)的转录水平显著降低(图1d-f);
综上说明MdRLKT1 通过激活寄主防御反应正向调控抗病性。
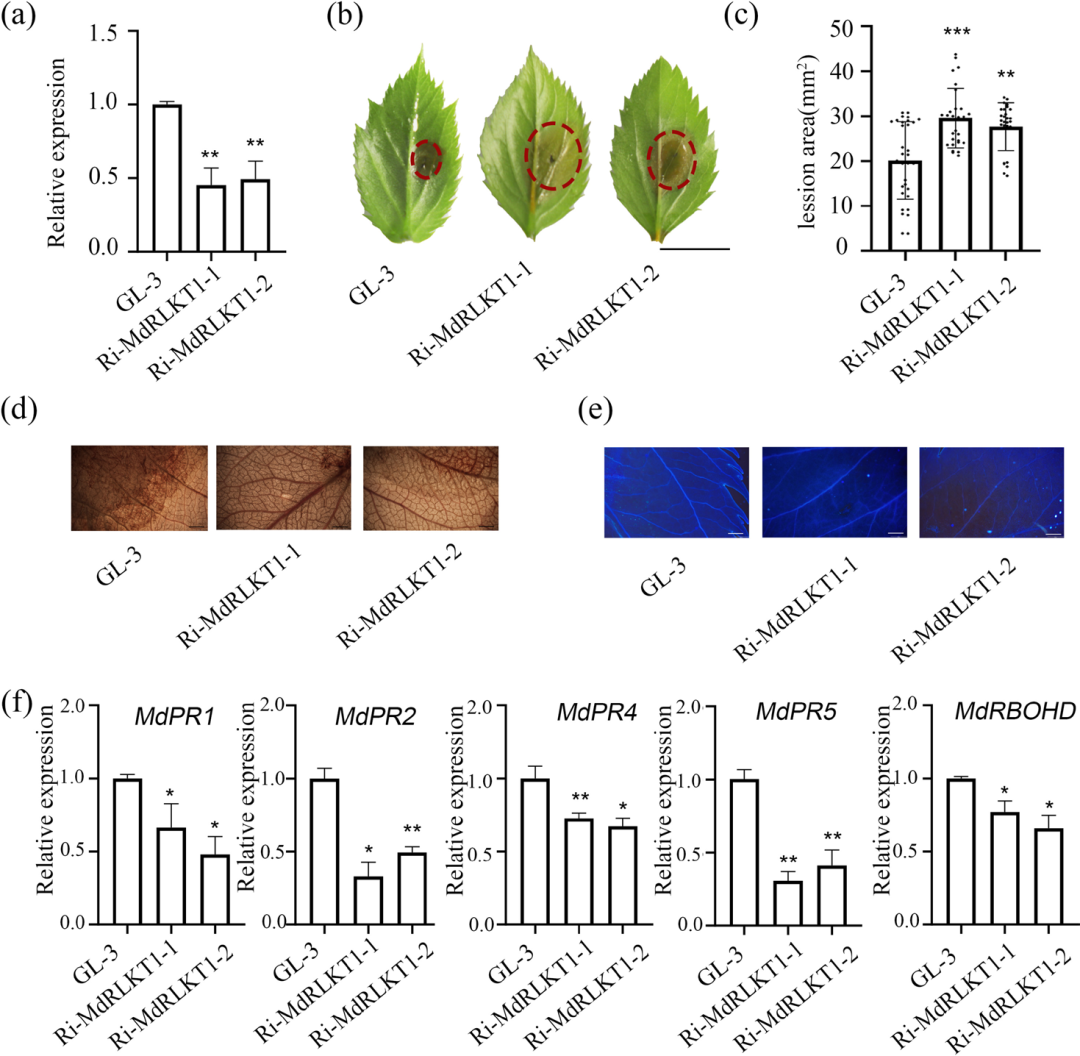
图1.MdRLKT1正向调控苹果对腐烂病菌(Cytospora mali )的抗性
2. MdRLKT1 与转录因子 MdRAX2 直接互作并促进其入核
为阐明MdRLKT1在植物免疫中的作用,本研究以MdRLKT1为诱饵,过酵母双杂交(Y2H)筛选发现 MdRLKT1 与 MYB 类转录因子 MdRAX2 互作,并通过 Co-IP、LCI、BiFC 和 MST 等多种实验在体内外验证了确实存在直接物理相互作用,且互作场所发生在细胞膜上;进一步通过亚细胞定位实验发现,MdRAX2 具有细胞核与质膜的双重定位特征。而当 MdRLKT1 与 MdRAX2 共表达时,MdRAX2 在细胞核中的积累显著增加,表明 MdRLKT1 促进了 MdRAX2 的入核转运(图2)。
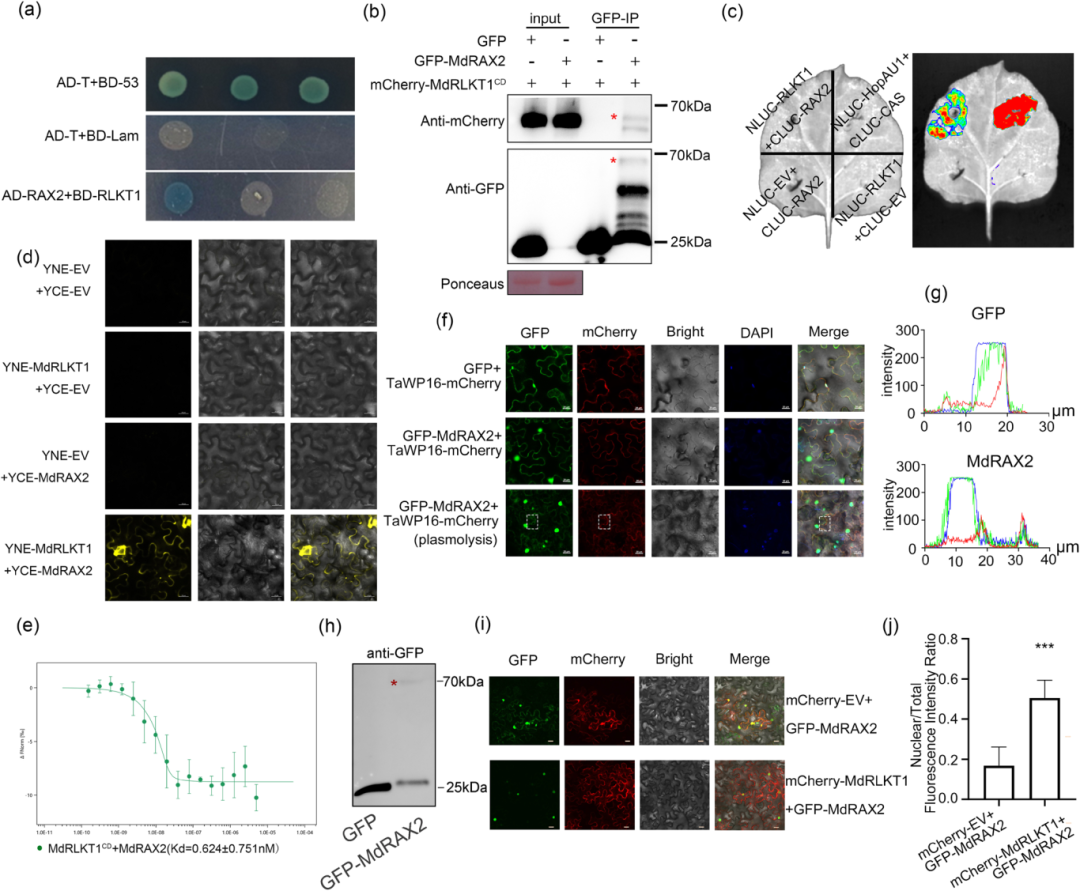
图2. MdRLKT1与MdRAX2相互作用并促进MdRAX2入核
3. MdRAX2 在苹果抗腐烂病菌中起正调控作用
为验证MdRAX2在苹果抗腐烂病菌中的作用,研究者在苹果叶片中进行瞬时表达实验,结果显示:
·正向抗病作用:瞬时过表达 MdRAX2 的苹果叶片对腐烂病菌的抗性显著增强,病斑面积比对照组缩小了 10.6%。此外,过表达植株在染病后表现出更强的活性氧积累和木质素沉积(图3a-e);
·沉默导致感病:利用 RNAi 技术干扰 MdRAX2 的表达后,叶片的病斑面积比对照组扩大了 41.8%,且 ROS 和木质素的积累明显减少(图3f-j);
·遗传互作逻辑:过表达 MdRLKT1 显著增强苹果叶片对 C. mali 的抗性;但在过表达 MdRLKT1 的背景下同时沉默 MdRAX2,则完全消除了 MdRLKT1 过表达所诱导的抗病表型,表明MdRAX2 是 MdRLKT1 介导苹果抗腐烂病菌所必需的。
结论:MdRAX2 是苹果免疫响应的正向调控因子,它通过促进防御物质(ROS 和木质素)的积累来增强苹果对腐烂病的抵抗力,且在 MdRLKT1 介导的防御通路中处于核心地位。
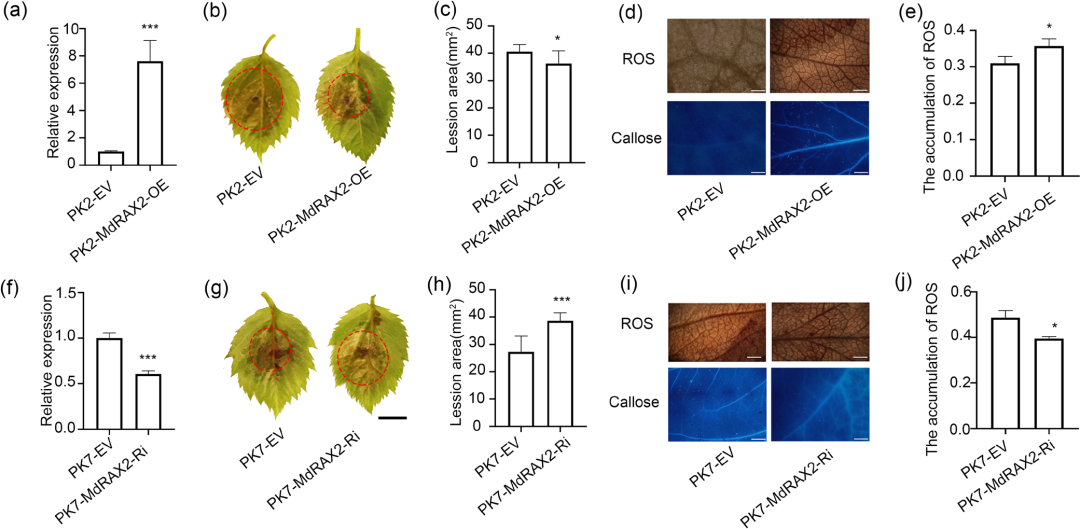
图3.MdRAX2正向调控苹果对腐烂病菌的抗性
4. MdRLKT1在体外磷酸化MdRAX2
为探究MdRAX2是否是MdRLKT1的磷酸化底物,在确认二者直接相互作用后,进一步进行了体外激酶活性检测。
磷酸化实验:体外激酶实验显示,MdRLKT1 能够磷酸化 MdRAX2,且这种磷酸化依赖于 MdRLKT1 的激酶活性。
位点鉴定:通过质谱(LC-MS/MS)和定点突变进一步确认,MdRAX2 的第 147 位丝氨酸(Ser147)是关键的磷酸化位点。
功能验证:突变体 MdRAX2(S147A)(模拟去磷酸化状态)失去了正常 MdRAX2 能够增强苹果抗性的功能,证明 Ser147 的磷酸化对于 MdRAX2 发挥抗病功能至关重要。
5. MdRAX2 结合 MdMKS1 的启动子并抑制其表达
由于MdRAX2具有转录因子功能,其可能通过调控靶基因表达参与免疫应答过程。本研究通过 DAP-seq、EMSA 和 Y1H 实验证实,转录因子 MdRAX2 直接结合在负调控基因 MdMKS1 启动子的 AG 富集基序上。LUC 和 GUS 活性分析进一步证明 MdRAX2 充当转录抑制子,显著下调 MdMKS1 的表达(图4)。
关键在于,这种抑制功能高度依赖其 Ser147 位点的磷酸化状态:S147A 突变虽不改变结合能力,但会导致其抑制活性完全丧失;而经受体激酶 MdRLKT1 磷酸化后,MdRAX2 对 MdMKS1 启动子的结合亲和力显著增强。这表明 MdRAX2 必须通过磷酸化触发的“开关”效应,才能精准关停免疫负调控因子,从而激活苹果的抗病响应。
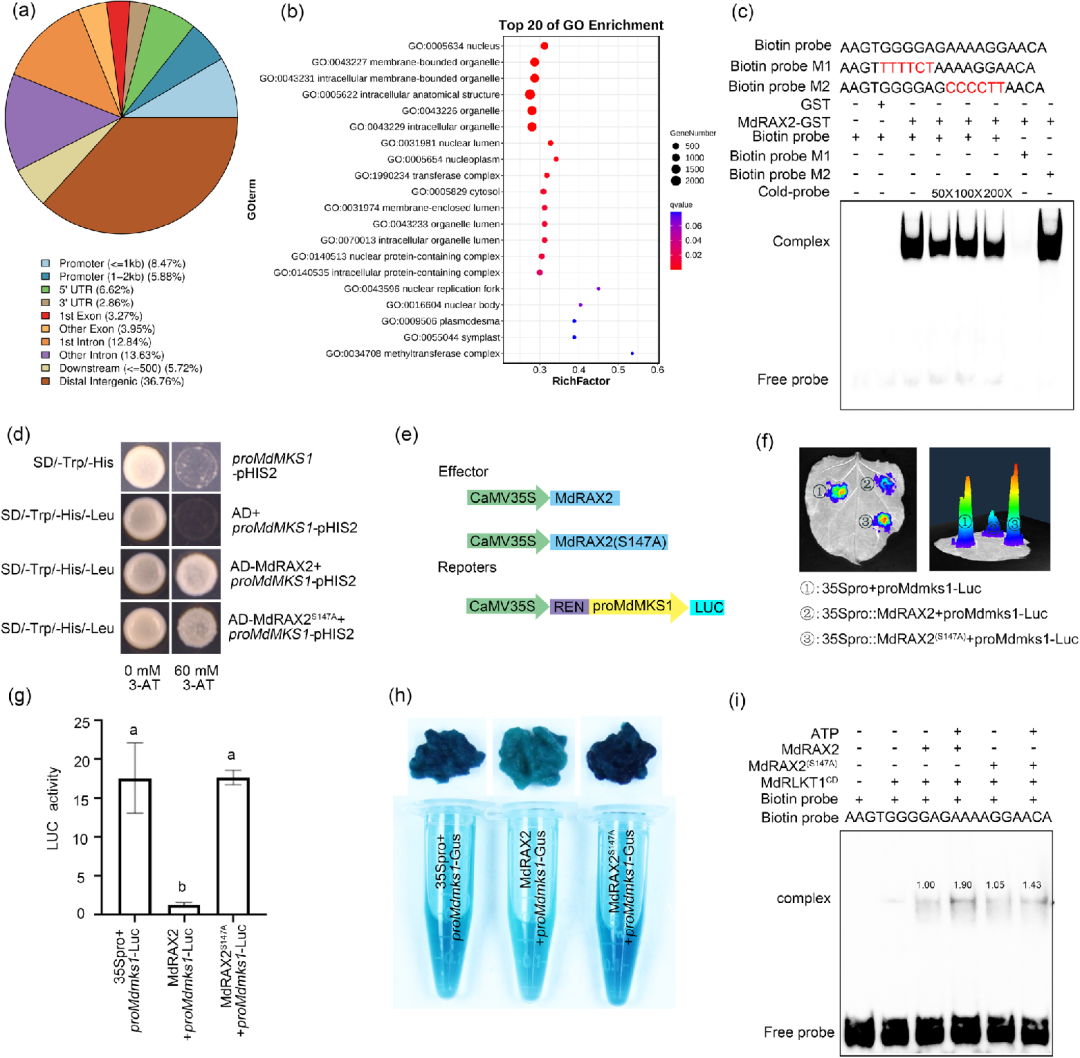
图4.MdRAX2与MdMKS1启动子结合并抑制其表达
6. MdMKS1 负向调节苹果对腐烂病菌的抗性
MKS1作为丝裂原活化蛋白激酶(MAPKs)的底物蛋白,是MAPK信号通路中的介导因子。为探究MdMKS1在苹果抗性中的作用,研究人员在苹果叶片中瞬时过表达或沉默该基因。结果显示瞬时过表达 MdMKS1 会显著削弱苹果的抗性并抑制防御相关基因(PR 基因)的表达;而沉默 MdMKS1 则能显著增强抗性,说明MdMKS1 在苹果免疫中充当“刹车”或负调控因子的角色。
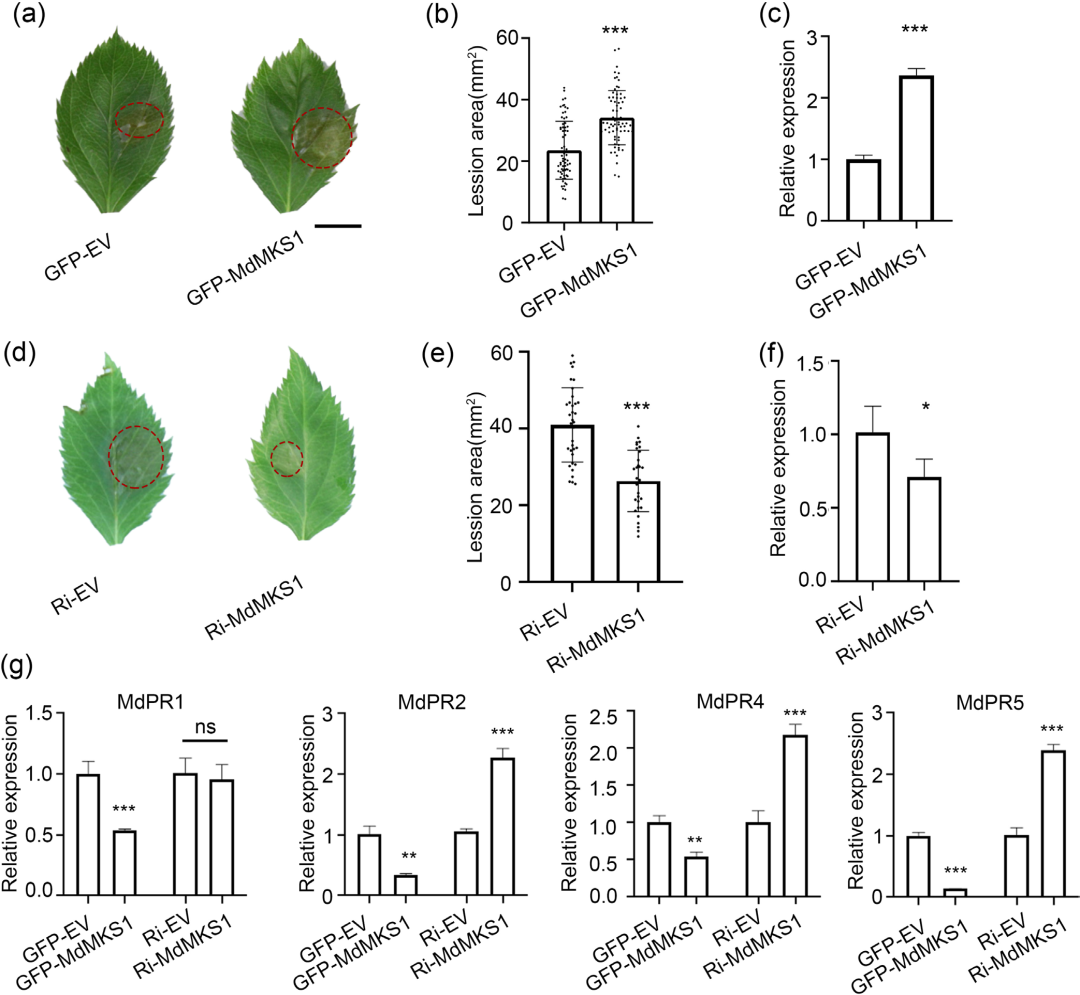
图5. MdMKS1通过抑制病原相关(PR)基因的表达,调节对苹果腐烂病菌的抗性
研究总结

图6. MdRLKT1–MdRAX2–MdMKS1级联调控苹果对C. mali的抗性模式图
该研究揭示了苹果抗腐烂病的免疫调控新机制:研究者利用酵母双杂交(Y2H)等技术筛选并证实了受体样激酶 MdRLKT1 与转录因子 MdRAX2 的直接互作。感病时,MdRLKT1 磷酸化 MdRAX2 促进其入核;结合DAP-seq、Y1H等实验证明了MdRAX2 直接结合并抑制负调控因子 MdMKS1 的表达,从而解除其对免疫系统的限制。该研究构建了由“激酶—转录因子—功能基因”组成的 MdRLKT1–MdRAX2–MdMKS1 调控模块,该磷酸化级联模块通过精准“关停”免疫刹车,显著增强了苹果的抗病性。
原文链接
https://doi.org/10.1111/pbi.70621
西北农林科技大学植物保护学院博士研究生唐亚楠为该论文第一作者,黄丽丽教授和冯浩教授为论文的通讯作者。作物抗逆与高效生产全国重点实验室实验平台为研究提供了技术支撑,该研究得到了国家自然科学基金项目的资助。